
Enllaç a la primera part.
Algunes de les estructures secundàries de l'aparell masticatori dels tireòfors que podrien tenir un paper rellevant en el processament dels aliments serien la ramfoteca o bec queratinós, un paladar secundari, les "galtes" i l'aparell hioïdal (els ossos de la llengua) i la forma del musell. La ramfoteca, a més de reduir l'estrès del crani tal com es va mostrar a la primera part, hauria estat una estructura molt eficaç a l'hora de tallar les plantes, potser fins i tot tant com les pròpies dents. Així doncs, el grau de desenvolupament de la ramfoteca estaria relacionat amb diferents estratègies de pastura, en cadascuna de les quals la duresa de les plantes que s'escollirien seria diferent. En els anquilosaures també s'hi ha identificat un paladar secundari més o menys desenvolupat que es troba format pels següents ossos: premaxil·la, maxil·lar, vòmer, palatí i pterigoide; la qual cosa indica que s'estendria des del principi de la mandíbula superior fins al darrera de la filera maxil·lar de dents. És possible que el paladar secundari facilités moviments masticatoris complexos perquè actuaria com una estructura de reforç de les mandíbules. Això sembla coherent amb el patró de mecanismes mandibulars identificat per l'equip d'en Attila Ősi l'any 2016, ja que els anquilosàurids presenten un paladar secundari més complex que els nodosàurids (Pereda-Suberbiola, 2007).
Fonts:
Algunes de les estructures secundàries de l'aparell masticatori dels tireòfors que podrien tenir un paper rellevant en el processament dels aliments serien la ramfoteca o bec queratinós, un paladar secundari, les "galtes" i l'aparell hioïdal (els ossos de la llengua) i la forma del musell. La ramfoteca, a més de reduir l'estrès del crani tal com es va mostrar a la primera part, hauria estat una estructura molt eficaç a l'hora de tallar les plantes, potser fins i tot tant com les pròpies dents. Així doncs, el grau de desenvolupament de la ramfoteca estaria relacionat amb diferents estratègies de pastura, en cadascuna de les quals la duresa de les plantes que s'escollirien seria diferent. En els anquilosaures també s'hi ha identificat un paladar secundari més o menys desenvolupat que es troba format pels següents ossos: premaxil·la, maxil·lar, vòmer, palatí i pterigoide; la qual cosa indica que s'estendria des del principi de la mandíbula superior fins al darrera de la filera maxil·lar de dents. És possible que el paladar secundari facilités moviments masticatoris complexos perquè actuaria com una estructura de reforç de les mandíbules. Això sembla coherent amb el patró de mecanismes mandibulars identificat per l'equip d'en Attila Ősi l'any 2016, ja que els anquilosàurids presenten un paladar secundari més complex que els nodosàurids (Pereda-Suberbiola, 2007).
 |
| Reconstrucció del cap i el coll de l'estegosaure del Juràssic Superior de Portugal Miragaia. La ramfoteca només s'aprecia a la mandíbula inferior en aquesta il·lustració perquè investigacions recents sobre els cranis dels estegosaures indiquen que no tindrien ramfoteca a la mandíbula superior. Foto: PrehistoryByLiam. |
 |
| Situació del paladar secundari al crani de l'Euoplocephalus. Foto: Bourke et al. (2018)/PLoS ONE. |
Se ha suggerit la presència de "galtes" en tireòfors i en altres dinosaures ornitisquis, les quals servirien per a impedir que el menjar caigués de la boca durant el seu processament, encara que en alguns altres clades d'ornitisquis, com els ceratopsians, aquesta funció alimentària ha estat descartada per bastants paleontòlegs. En el cas dels estegosaures, aquest paper ja el desenvoluparia la làmina dorsal del dentari que ja es va mencionar a la primera part. Això no sols indica que els estegosaures no tindrien "galtes", si no també l'absència d'una ramfoteca allargada que podria fer una funció semblant a part de tallar les plantes (la qual també existiria només a la mandíbula inferior tal com es veu a la reconstrucció del Miragaia). On sí que s'hi han identificat estructures semblants a les galtes dels mamífers és en els nodosàurids, els quals mostren osteoderms de forma oval a la zona equivalent a la galta mamífera. Aquests osteoderms no estan articulats ni a les mandíbules inferiors ni als maxil·lars, la qual cosa és coherent amb un moviment limitat de les "galtes" que hauria permès que l'aliment no s'empassés sense mastegar mentre el processaven les dents. En els anquilosàurids, aquesta estructura o no s'identifica o té una extensió molt menor que en els nodosàurids, la qual cosa mostra que no la necessitarien de la mateixa manera que en els nodosàurids (Pereda-Suberbiola, 2007; Ősi et al., 2016).
 |
| Cranis complets de Panoplosaurus (esquerra) i Edmontonia (dreta) en vista lateral. L'abreviació cp es refereix als osteoderms de les "galtes". Foto: Ősi et al. (2016)/Historical Biology. |
L'existència d'un aparell hioïdal ben desenvolupat en els dinosaures no-aviaris en general i en els tireòfors en particular, un element que s'observa en totes les aus i els rèptils actuals, podria suggerir la presència d'una llengua llarga i flexible que els ajudaria a recollir i introduir les plantes a la boca i a mantenir-les en aquesta durant el procés masticatori. La primera referència sobre la conservació d'un aparell hioïdal complet en un tireòfor es va publicar l'any 2015 gràcies a un equip d'investigació liderat per en Robert Hill (Facultat de Medicina Osteopàtica de Nova York) i l'estructura en qüestió es va conservar en un esquelet parcial de Pinacosaurus, anquilosàurid del Cretaci Superior Campanià de Mongòlia, procedent del jaciment d'Ukhaa Tolgod (Formació Djadokhta). L'aparell hioïdal del Pinacosaurus està compost per dues paraglossalia, dos ossos de forma triangular que fins llavors només s'havien identificat en aus; dos primers ceratobranquials, dos ossos en forma de barres aplanades i que es troben darrere de les paraglossalia; dos segons ceratobranquials ossos en forma de canya que es troben darrere dels primers ceratobranquials i dos epibranquials, uns altres ossos allargats però més petits que els altres que es van trobar desarticulats però que per comparació amb les aus i els rèptils actuals es situarien darrere dels segons ceratobranquials. Als segons ceratobranquials hi ha una superfície articular de forma oval que es troba a l'extrem anterior de l'os, la qual cosa indica que aquests ossos s'ajuntaven amb una estructura cartilaginosa al mig de l'aparell hioïdal que es correspondria amb el basihial de les aus i els rèptils actuals. Gràcies a aquest descobriment en el Pinacosaurus, es van poder identificar ossos de l'aparell hioïdal, entre ells les paraglossalia, en altres anquilosàurids (Saichania i Euoplocephalus) i també al nodosàurid Edmontonia. Per tant, es va confirmar la presència d'una llengua llarga i musculosa en els anquilosaures. I no només això, si no que la presència de les paraglossalia en dinosaures tant distants entre sí com els anquilosaures i les aus indica que aquests ossos formarien part de l'aparell hioïdal de tots els dinosaures des del seu origen, estant absents en el registre fòssil d'altres llinatges perquè tindrien una constitució cartilaginosa o simplement perquè no s'han pogut identificar com a tals fins ara. Les evidències de l'aparell hioïdal en dinosaures que no siguin ni anquilosaures ni aus només consisteixen en primers i segons ceratobranquials (Pereda-Suberbiola, 2007; Hill et al., 2015).
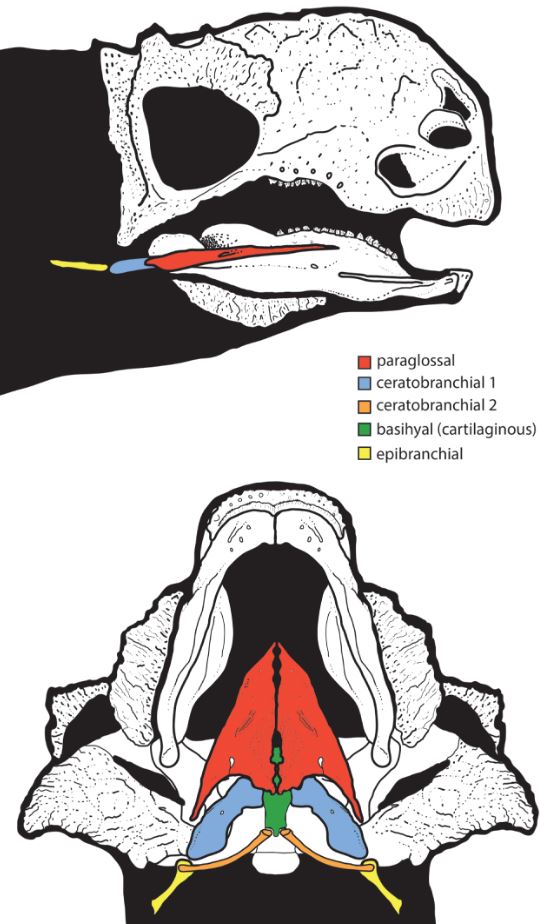 |
| Reconstrucció de l'aparell hioïdal del Pinacosaurus amb diferenciació dels seus elements. Foto: Hill et al. (2015)/Zoological Journal of the Linnean Society. |
En els mamífers, s'ha relacionat la forma del musell de les espècies herbívores amb el tipus de vegetals dels que s'alimenten. Així doncs, aquells mamífers que s'alimenten de les fulles dels arbres, com ara els girafes i els okapis, tenen musell estrets, els que s'alimenten d'herba, com els nyus, tenen musells amples i els que s'alimenten a la vegada d'herba i de fulles, com els impales, tenen un musell més estret que els dels nyus però més ample que el de les girafes. Malgrat que aquesta diferenciació utilitzada per als mamífers no es pot acabar d'aplicar als dinosaures no-aviaris perquè els prats d'herba no existien al Mesozoic, un fenomen semblant es dóna entre aquelles espècies de tireòfors amb uns musells més estrets o més amples, els quals es correspondrien de forma respectiva amb animals d'un comportament alimentari selectiu i animals d'un comportament alimentari generalista (la qual cosa no acaba de ser del tot diferent del que passa amb els mamífers actuals ja que les espècies que s'alimenten d'herba solen triar les parts més nutritives de la planta mentre que els que basen la seva dieta en les fulles d'arbres no discriminen tant a l'hora d'elegir l'aliment). El musell és bastant estrets en bona part dels estegosaures, mentre que en els anquilosaures s'observa una ampliació de la mida del musell entre les espècies primitives tant de nodosàurids com d'anquilosàurids del Juràssic Superior i el Cretaci Inferior i les derivades de cada clade del Cretaci Superior. Per tant, els nodosàurids i els anquilosàurids van desenvolupar de forma independent la capacitat de menjar a partir d'una major varietat i quantitat de matèria vegetal a mitjans del Cretaci (Pereda-Suberbiola, 2007; Ősi et al., 2016).
 |
| Comparació entre els musells de diferents nodosàurids (A-G) i anquilosàurids (H-L). Es pot comprovar que en les últimes espècies de cada sèrie el musell és més ample que en les primeres. Foto: Ősi et al. (2016)/Historical Biology. |
Tenint en compte que els tireòfors eren en la seva majoria animals quadrúpedes obligats de no gaire altura, és probable que la major part del seu repertori alimentari, sobretot durant el Juràssic i la primera meitat del Cretaci, estaria basat en ciques, bennettitals i petites coníferes, així com en plantes de l'ordre Caytoniales, unes pteridospermes o "falgueres amb llavors" pròpies del Mesozoic que en alguns casos s'han considerat avantpassades de les angiospermes. La presència d'un moviment mandibular palinal en els nodosàurids i els anquilosàurids més derivats suggereix que estaven adaptats a alimentar-se de fulles, tiges i llavors que contenien una proporció de cel·lulosa important i que, per tant, necessitaven ser processats amb suficient precisió a la boca abans de passar a l'estómac. Tenint en compte que estem parlant del Cretaci Superior, entre aquestes plantes es troben angiospermes de sotabosc com les representades per les famílies Araceae (com les llenties d'aigua) i Proteaceae (com la macadàmia). Curiosament, aquest moviment mandibular palinal només s'ha localitzat en els nodosàurids i anquilosàurids derivats d'Amèrica del Nord i Europa, la qual cosa es correspon amb el clima humit que tindrien aquestes regions durant el Cretaci Superior, factor que afavoreix l'aparició de plantes dures. Per contra, l'absència del moviment palinal en els nodosàurids i anquilosàurids derivats del Cretaci Superior d'Àsia (i la presència, en el seu lloc, d'un moviment ortal més simple) es relaciona amb el clima àrid existent en aquesta conjuntura espaciotemporal, la qual cosa implica l'existència de plantes menys dures que en els climes més humits i que, en conseqüència, no necessitarien un processament digestiu tant meticulós. L'engrandiment de l'amplada del musell en els membres més derivats de cada família d'anquilosaures podria tractar-se d'una adaptació al consum d'angiospermes a causa que aquesta novetat evolutiva apareix quan les plantes amb flors es converteixen en les plantes dominants, encara que aquesta hipòtesi no està demostrada en aquest moment (Pereda-Suberbiola, 2007; Ősi et al., 2016).
 |
| Fulla fòssil de Sagenopteris, una representant de Caytoniales del Triàsic, el Juràssic i el Cretaci Inferior. Foto: Abbieeturner. |
Encara que les plantes serien l'element principal de la dieta dels tireòfors, també s'ha suggerit al llarg de dècades que els "dinosaures cuirassats" completarien la seva dieta amb la ingesta d'animals, una hipòtesi que en els últims anys s'ha pogut comprovar per a alguns casos en particular. La primera suposició sobre una dieta "carnívora" és del paleontòleg hongarès Franz Nopcsa i data del 1928, la qual diu que el Scolosaurus, un anquilosàurid del Cretaci Superior Campanià del Canadà, hauria subsistit de la ingesta d'un 1 kg d'insectes al dia com a adaptació a un ambient desèrtic. Una visió contrària a la d'en Nopcsa és la d'en Kenneth Carpenter, que va rebatre aquesta idea d'un anquilosàurid insectívor argumentant que els animals endoterms actuals que subsisteixen exclusivament dels insectes no superen els 30 kg de pes, per la qual cosa seria molt difícil la supervivència d'un dinosaure d'un pes al voltant de la tona com era el Scolosaurus només amb insectes. El 1977, la paleontòloga polonesa Teresa Maryańska va proposar que els anquilosàurids serien carronyaires ocasionals a causa de la mida petita de les dents en relació amb el crani i el desenvolupament escàs de l'esmalt dental, però és una idea que ha rebut poc suport entre altres paleontòlegs. La descoberta de l'aparell hioïdal complet del Pinacosaurus el 2015 va servir perquè la idea d'un tirèofor insectívor d'en Nopcsa tornés a tenir validesa ja que la llengua musculosa d'aquest dinosaure seria semblant a la de molts animals insectívors actuals. Tenint en compte que a Ukhaa Tolgod els fòssils vegetals no són gaire comuns però sí que ho són els d'insectes, la idea d'una alimentació parcialment insectívora seria viable per al Pinacosaurus, el qual, amb uns 5 m de llargada i 1,9 t de pes, seria un anquilosàurid relativament petit (Pereda-Suberbiola, 2007; Hill et al., 2015).
 |
| Icnofòssils d'escarabats (Fictovichnus) procedents d'Ukhaa Tolgod. Els insectes productors d'aquestes estructures serien les preses potencials del Pinacosaurus. Foto: Johnston et al. (1996)/Canadian Journal of Earth Science. |
Encara que al final la força mandibular dels estegosaures s'ha demostrat més potent del que sempre s'ha pensat, alguns paleontòlegs també han apuntat que aquests dinosaures van adoptar estratègies alimentàries pròpies que els van permetre ser un dels grups de dinosaures vegetarians més abundants del Juràssic Mitjà i Superior. A causa que els estegosaures tenen les extremitats posteriors més llargues que les anteriors, en Robert Bakker va apuntar l'any 1986 que es recolzarien sobre les seves potes del darrere i també la seva cua per a poder arribar a una vegetació situada a una altura a la qual no podrien arribar en una posició quadrúpeda. Però aquesta hipòtesi es pot contraaargumentar en dos sentits. El primer és que seria difícil per als estegosaures adoptar una postura bípeda recolzant-se sobre la seva cua a causa que aquesta era una part del cos molt rígida a causa de tota la sèrie de plaques que presentava. El segon és fent una comparació amb un mamífer herbívor de gran mida com és l'elefant, el qual pot arribar a les copes dels arbres a través de l'adopció d'una postura bípeda temporal però que aquesta no és l'habitual quan s'alimenten (Pereda-Suberbiola, 2007).
 |
| Reconstrucció de l'esquelet de l'estegosaure del Juràssic Superior de la Xina Gigantspinosaurus exposada al Museu de Dinosaures de Zigong (Xina), en la qual el dinosaure mostra una postura bípeda. Foto: David Stanley. |
L'oclusió dental és molt escassa o inexistent en el Scutellosaurus i l'Emausaurus, considerats tradicionalment els tireòfors més basals. Aquesta és una condició semblant a la del possible tireòfor basal del Triàsic Lesothosaurus, per la qual cosa seria la condició primitiva del contacte interdental en aquest clade de dinosaures. Aquesta situació es continua observant en alguns estegosaures. Per contra, el mecanisme mandibular del Scelidosaurus s'assembla a la d'alguns anquilosaures més basals, la qual cosa es podria interpretar com l'evidència d'un parentesc més pròxim entre el Scelidosaurus i els anquilosaures del que s'havia pensat, que el mecanisme ortal de punció-trituració va aparèixer independentment en el Scelidosaurus i els anquilosaures o que els estegosaures van tornar a la condició primitiva a partir de la més moderna. En els anquilosaures més basals, s'observen tot un seguit de caràcters que es perden (les dents premaxil·lars) o que es modifiquen bastant (un cíngol molt poc desenvolupat). En els nodosàuris europeus derivats es retenen les dents premaxil·lars i un musell estret en comparació amb els seus equivalents nord-americans, encara que l'absència de nodosàurids europeus amb musells amples i premaxil·les desdentades correspondria més a un biaix de la preservació de fòssils més que no pas a una autèntica falta d'aquests animals en l'ecosistema. Una diferència substancial entre les mandíbules inferiors dels nodosàurids i les dels anquilosàurids és la presència en els primers d'un procés coronoide (l'extensió de la mandíbula que serveix d'inserció dels músculs que la mouen) bastant més desenvolupat que el dels darrers, la qual cosa seria una evidència que hi hauria certes diferències en la mida i l'angle d'inserció dels músculs mandibulars entre els dos clades de dinosaures que farien que els nodosàurids tinguessin una força de mossegada major que la dels anquilosàurids (Pereda-Suberbiola, 2007; Ősi et al., 2016).
 |
| Cladograma simplificat i calibrat temporalment dels tireòfors en el qual s'indiquen les diferents innovacions en l'aspecte mandibular a través de dibuixos i diferents colors en les línies corresponents a cada tàxon. Foto: Ősi et al. (2016)/Historical Biology. |
Hill, R. V., D'Emic, M. D., Bever, G. S. & Norell, M. A. (2015). "A complex hyobranchial apparatus in a Cretaceous dinosaur and the antiquity of avian paraglossalia." Zoological Journal of the Linnean Society, Vol. 175 (4): p. 892-909. https://doi.org/10.1111/zoj.12293
Ősi, A., Prondvai, E., Mallon, J. & Bodor, E. R. (2016). "Diversity and convergences in the evolution of feeding adaptations in ankylosaurs (Dinosauria: Ornitischia)." Historical Biology, Vol. 29 (4): p. 539-570. https://doi.org/10.1080/08912963.2016.1208194
Pereda-Suberbiola, X. (2007). "Dieta y modo de alimentación de los dinosaurios acorazados." Dins de: Sanz, J. L. (ed.), Los dinosaurios en el siglo XXI, p. 187-207. Tusquets Editores. ISBN: 978-02-5333-497-8.
Cap comentari:
Publica un comentari a l'entrada